James Mallet
University College London
Preprint of article in "Encyclopedia of Biodiversity",
ed. Simon A. Levin
Academic Press, New York. (2001) Volume 5: pp.
427-440.
I. What are species concepts for?
II. Statement of bias
III. Darwinian species criteria
A. Darwin's species criterion
B. Polytypic species
IV. The philosophization of species: the interbreeding species concept
V. Alternative species concepts
A. Ecological species concept
B. Recognition concept of species
VI. Species concepts based on history
A. Monophyly
B. Genealogy
C. Diagnostic species concept
VII. Combined species concepts
A. Evolutionary and lineage concepts
B. Cohesion concept of species
VIII. Dissent: maybe species are not real
A. Taxonomic practice
B. Populations are the evolutionary units,
not species
C. Phenetic species concept
D. Genotypic cluster criterion
E. The unreality of species in space and time
IX. The importance of species concepts for biodiversity and conservation
A. Traditional: species as real entities
B. Alternative: genetic differences more important
than species status
C. Species differences as ecologically important
markers
D. Biodiversity in space and time
SPECIES ARE CRUCIAL IN MANY BIODIVERSITY ISSUES: much of conservation, biodiversity studies, ecology, and legislation concerns this taxonomic level. It may therefore seem rather surprising that biologists have failed to agree on a single species concept. The disagreement means that species counts could easily differ by an order of magnitude or more when the same data is examined by different taxonomies. This article explores the controversy on species concepts and its implications for evolution and conservation.
I. What are species concepts for?
Individual organisms can usually be recognized, but the larger units we use to describe the diversity of life, such as populations, subspecies, or species are not so easily identifiable. Taxonomists further group species into genera, families, orders, and kingdoms etc., while ecologists group species into higher structures such as communities and ecosystems. The justification for these group terms is utility, rather than intrinsic naturalness, but as far as possible we attempt to delimit groups of organisms along natural fault lines, so that approximately the same groupings can be recovered by independent observers. However there will be a virtually infinite number of different, albeit nested, ways of classifying the same organisms, given that life has evolved hierarchically.
Darwin (1859) felt that species were similar in kind to groupings at lower and higher taxonomic levels; in contrast, most recent authors suggest that species are more objectively identifiable, and thus more "real" than, say, populations or genera. Much of ecology and biodiversity today appears to depend on the idea that the species is the fundamental taxon, and these fields could be undermined if, say, genera, or subspecies had the same logical status.
Species concepts originate in taxonomy, where the species is "the basic rank of classification" according to the International Commission of Zoological Nomenclature. The main use of species in taxonomy and derivative sciences is to order and retrieve information on individual specimens in collections or data banks. In evolution, we would like to delimit a particular kind of evolution, speciation, which produces a result qualitatively different from within-population evolution, although it may of course involve the same processes. In ecology, the species is a group of individuals within which variation can often be ignored for the purposes of studying local populations or communities, so that species can compete, for example, while subspecies or genera are not usually considered in this light. In biodiversity and conservation studies, and in environmental legislation, species are important as units which we would like to be able to count both regionally or globally.
It would be enormously helpful if a single definition of species could satisfy all these uses, but a generally accepted definition has yet to be found, and indeed is believed by some to be an impossibility. A unitary definition should be possible, however, if species are "real", objectively definable and fundamental biological units. Conversely, even if species have no greater objectivity than other taxa, unitary nominalistic guidelines for definining species might still be found, perhaps after much diplomacy, via international agreement among biologists; after all, if we can adopt metres and kilograms, we should be able to agree on units of biodiversity in a similar way. In either case, knowledge of the full gamut of today's competing solutions to the species concept problem will probably be necessary for a universal species definition to be found. This article reviews the proposals currently on the table, and their usefulness in ecology, evolution, and conservation.
II. Statement of bias
I am of the opinion that the "reality" of species in evolution, and in ecological and biodiversity studies over large areas has been overestimated. In contrast, it is clear to any naturalist that species are usually somewhat objectively definable in local communities. It is my belief that confusion over species concepts has been caused by scientists not only attempting to extend this local objectivity of species over space and evolutionary time, but also arguing fruitlessly among themselves as to the nature of the important reality that underlies this illusory spatiotemporal objectivity. To me, agreement on a unified species-level taxonomy is possible, but will be forthcoming only if we accept that species lack a single, interpretable biological reality over their geographic range and across geological time.
Just as Marxist theory may be wrong, yet remains a convenient tool for studying political history, I hope that my own views can provide, even for the skeptic, a useful framework on which the history of proposals for species concepts can be compared. A variety of other outlooks can be found in Mayr (1982), Cracraft (1989), Ridley (1996), Claridge et al. (1997) and Howard & Berlocher (1998).
III. Darwinian species criteria
A. Darwin's morphological species criterion
Before Darwin, it was often assumed that each species had an Aristotelian "form" or "essence", and that variation within a species was due to imperfections in the actualization of this form. Each individual species was defined by its essence, which itself was unvarying and inherently different from all other species essences. This mode of thought of course precluded transformation of one species into another, and was associated with belief that each form was separately created by God. Darwin's extensive travels and knowledge of taxonomy led to a realization that the distinction between intraspecific and interspecific variation was false. His abandonment of the essentialist philosophy and its species concept went hand in hand with his appreciation that variation itself was among the most important characteristics of living organisms, because it was this variation which allowed species to evolve.
Darwin guessed (correctly) that essentialist species would be hard to give up: "... we shall have to treat species in the same manner as those naturalists treat genera, who admit that genera are merely artificial combinations made for convenience. This may not be a cheering prospect; but we shall at least be freed from the vain search for the undiscovered and undiscoverable essence of the term species." (Darwin 1859: 485). He argued that species were little more than varieties which acquired their claim to a greater reality only when intermediates died out leaving a morphological gap: "... I believe that species come to be tolerably well-defined objects, and do not at any one period present an inextricable chaos of varying and intermediate links" (Darwin 1859: 177). This morphological gap criterion, which seems to have been accepted by most early evolutionists (e.g. Wallace 1865, Robson 1928), has been called a "morphological species concept" because Darwin used the gaps in morphology to delimit species; however, it would be easy to extend his species criterion to ecology, behaviour, or genetic (see VIII D below).
B. Polytypic species
A major revolution in zoological taxonomy occurred around 1900. As the great museum collections became more complete, it became obvious that apparently distinct species found in different areas frequently intergraded where they overlapped. These replacement species were usually combined as subspecies within a "polytypic" species (see SUBSPECIES, SEMISPECIES, SUPERSPECIES), an idea suggested for "geographical varieties" by early systematists and Darwinists such as Wallace (1865). The taxonomic clarification that followed, which allowed identifiable geographic varieties to be named below the species level as subspecies, was conceptually more or less complete by the 1920s and 1930s. At the same time, other infraspecific animal taxa such as local varieties or forms were deemed unnameable in the Linnean taxonomy. These changes are now incorporated into the International Code of Zoological Nomenclature (see also SUBSPECIES, SEMISPECIES, SUPERSPECIES). Similar ideas were promoted in botany by G.L. Stebbins (see Mayr 1982), although local varieties and polymorphic forms remain valid and nameable taxa in the International Botanical Code.
IV. The philosophization of species, the "interbreeding" concept.
In January 1904, E.B. Poulton read his famous presidential address - "What is a species?" - to the Entomological Society in London. Following up some ideas raised (but immediately dismissed) by Wallace (1865), Poulton proposed "syngamy" (i.e. interbreeding) as the true meaning of species. Poulton and Wallace were both particularly knowledgeable about swallowtail butterflies (Papilionidae). In swallowtails, there were strong sexual dimorphisms: the female colour pattern often mimicks unrelated unpalatable butterflies while the male is non-mimetic. The females themselves are often polymorphic, each female form mimicking a different distasteful model. Under a morphological criterion each form could be designated as a different species, whereas mating observations in the wild showed that they were part of the same interbreeding group. Similar ideas were promoted by the botanist J.P. Lotsy, who termed the interbreeding species a "syngameon". In the 1930s T. Dobzhansky studied morphologically indistinguishable sibling species of Drosophila fruit-flies and concluded that Lotsy's approach had some value. A species will rarely if ever interbreed with its sibling; each chooses mates from within its own species. Dobzhansky proposed his own interbreeding species concept, later popularized by Mayr as the "biological species concept", so named because interbreeding was considered the single true biological meaning, or reality of the term species (reviewed by Mayr 1970, 1982).
A short definition of the biological species concept is: "Species are groups of interbreeding natural populations that are reproductively isolated from other such groups" (Mayr 1970). This concept was not so much new as a clarification of two distinct threads: (1) a local component, the Poulton/Dobzhansky interbreeding concept, and (2) a global component which extended the interbreeding concept to cover geographical replacement series of actually or potentially interbreeding subspecies (Mayr 1970), as in the pre-existing idea of polytypic species (see also SUBSPECIES, SEMISPECIES, SUPERSPECIES).
This extended interbreeding concept was, until recently, almost universally adopted by evolutionists. The species concept problem appeared to have been solved; species were interbreeding communities, each of which formed a gene pool reproductively incompatible with other such communities. The new concept answered both perceived problems of Darwin's morphological approach: (1) that under morphological criteria, distinct mutants and polymorphic variants within populations might be considered separate species, and (2) that sibling species might be misclassified morphologically as members of the same species. The new approach was promoted in a long series of books and articles by Dobzhansky, Mayr, and their followers. Mayr in particular was highly influential by justifying the taxonomic application of the polytypic species criteria in terms of the new concept of gene flow.
In order to adopt this change, it was necessary to see species in a new post-Darwinian light. Instead of species being defined simply, using man-made criteria based on demonstrable characters such as morphology, species became defined by characteristics important in their own maintenance, that is by means of their biological function (Mayr 1982). Significantly, the philosophical term "concept" came into vogue along with these ideas about species, and that the term "species problem", which hitherto referred to the problem of how species arose (Robson 1928), became instead the problem of defining what species were. The important features of species defined by the "biological concept" were that they were protected from gene flow by what Dobzhansky termed "isolating mechanisms", including prezygotic factors (ecological, mate choice and fertilization incompatibilities) and postzygotic factors (hybrid inviability and sterility caused by genomic incompatibilities). By going beyond simple character-based identification of species, the concept could not apply to all living things; for example, Dobzhansky simply concluded that asexuals (between which no interbreeding is possible) could not have species.
Poulton, Mayr and Dobzhansky emphasized that their new concept was based on the reality that underlay species, rather than being merely a criterion useful in taxonomy; in this new philosophical approach, taxonomic criteria and conceptual issues of species became separate, with taxonomic criteria taking a more minor role. The concept was true from first principles, and was therefore untestable: difficulties such as hybridization, intermediates, or inapplicability to many plants and asexuals caused taxonomic problems, but did not disprove or even challenge the underlying truth of the concept. These imperfect actualizations of species' true reality were expected in nature. Mayr claimed that the biological concept would do away with "typology" (his term for species definitions based on a fixed, unvarying type or Aristotelian essence), but in many ways it can be seen that the biological concept reverts to a new kind of essentialism, where evolutionary maintenance via interbreeding is the underlying reality or essence of species.
V. Alternative species concepts
It is interesting that exactly this kind of search for the essence of species had been criticized by Darwin (1859). In his chapter "Hybridism", he specifically argued against using hybrid sterility and zygote inviability as a cut and dried characteristic of species (although he made little mention of pre-mating factors). Oddly, Mayr (1982:269) claimed both that Darwin treated species "purely typologically [i.e. as an essentialist] as characterized by degree of difference", and also that Darwin "had strong, even though perhaps unconscious, motivation ... to demonstrate that species lack the constancy and distinctiveness claimed for them by the creationists." Whether it is reasonable to criticise Darwin in such a contradictory way can be debated, but it is clear that Mayr's proposition that interbreeding is the true essence or reality of species immediately laid itself open to debate. Although the interbreeding concept had a long run (and still does), proposals for different kinds of biological reality of species were eventually forthcoming. By proposing a single reality for species, Poulton, Dobzhansky, and Mayr opened the Pandora's box of alternative essences deemed more important by other biologists.
A. Ecological species concept
Asexual organisms such as the bdelloid rotifers can clearly be clustered into groups recognizable as taxonomic species (Hutchinson 1968). On the other hand, distinct forms such as oaks (Quercus), between which there are high rates of hybridization, can remain recognizably distinct even where they co-occur. This suggested to L. van Valen and others that the true meaning of species was occupancy of an ecological niche rather than interbreeding. This ecological idea became known as the "ecological species concept". It became clear to Mayr during the 1970s also (see Mayr 1982) that gene flow could not unite every population in a polytypic, biological species' range, and that stabilization of phenotype might be effected by ecologically mediated stabilizing selection (see also VII B and VIII B below) rather than purely because of gene flow.
B. Recognition concept of species
An important attack on the biological species concept came from H.E.H. Paterson in the early 1980s. His claims were twofold: first, the Dobzhansky/Mayr term "isolating mechanisms" implied reproductive isolation was adaptive, which Paterson felt was unlikely; second, the true reality underlying species was proposed to be prezygotic compatibility, consisting of mating signals and fertilization signals. According to Paterson (1985), this compatibility is strongly conserved by stabilizing selection, whereas isolating mechanisms such as hybrid sterility or inviability are non-adaptive and can be argued to be a result rather than a cause of species separateness. To Paterson the true reality of species must be adaptive. He termed his idea of species the "recognition concept" versus Mayr's "isolation concept", and its important characteristics "specific mate recognition systems" (SMRS) instead of isolating mechanisms. Species were defined as "that most inclusive population of individual biparental organisms which share a common fertilization system" (Paterson 1985).
The idea is generally recognized as a useful critique and has gained strong currency in some circles. However, it has been pointed out that SMRS are more or less the inverse of prezygotic isolating mechanisms, and that the recognition concept therefore differs from the biological species concept mainly by focussing on a subset of isolating mechanisms used within the interbreeding concept. The interbreeding concept had always stressed a common gene pool and compatibility within a species, as well as isolation between species.
VI. Species concepts based on history
A. Monophyly
The rise of cladistic methods revolutionised systematics by proposing that all classification should be based on the idea of monophyly. This new system formalized the principle that paraphyletic and polyphyletic taxa were unnatural groupings which should not be used in taxonomy. It was natural to attempt to apply this idea throughout systematics, all the way down to the species level, leading to a monophyly criterion of species, a type of "phylogenetic species concept" (Hennig 1968, see also the diagnostic definition below). Species were seen as forming when a single interbreeding population split into two branches or lineages that did not exchange genetic material. In a somewhat different formulation, the "cladistic species concept", species are branch segments in the phylogeny, with every branching event leading to a new pair of species (Ridley 1996). Otherwise, if only one of the two branches were recognized as new, the other branch would become paraphyletic.
Perhaps the main criticism of this idea is that it could, if applied in taxonomy, cause great nomenclatural instability. Monophyly exhibits fractal self-similarity and can exist at very high or very low levels of the phylogeny, so the precise level at which species taxa exist becomes unclear. Supposing a new monophyletic form is discovered overlapping with, but remaining distinct from, a closely related local form in the terminal branches of an existing species. Recognition of this taxon as a species would leave the remaining branches within the original species paraphyletic. Many other branch segments would then need to be recognized at the species level, even if they interbreed and have reticulate, intermingling phylogenies. Many phylogenetic systematists therefore adopt a different phylogenetic concept, the diagnostic concept (see below), which can allow paraphyly at the species level.
B. Genealogy
Another problem with a monophyly concept is that a single, true phylogeny of taxa may rarely exist: an organismal phylogeny is in fact an abstraction of the actual genetic history, consisting of multiple gene genealogies, some of which may undergo genetic exchange with other taxa. There is now good evidence that occasional horizontal gene transfer and hybridization may selectively transfer genetic material between unrelated forms. Furthermore there are multiple gene lineages within any population, so that, if such a population were to become geographically or genetically split into two distinct forms, it would be some time before each branch became fixed for different, reciprocally monophyletic gene lineages at any single gene. The idea of monophyly for whole genomes then becomes hard to define, especially near the species boundary. However annoying, phylogenetic methods and evolutionary theory must face up to these facts (Avise & Ball 1990, Maddison 1997). It has therefore been suggested that species should be defined when a consensus between multiple gene genealogies indicates reciprocal monophyly. This is called the "genealogical species concept" (Baum & Shaw 1995).
Critics argue that this idea has many problems in common with other monophyly concepts of species (Davis 1997). Geographic forms that have become isolated in small populations or on islands, say, could rapidly become fixed for gene lineages, and become viewed as separate species without any biologically important evolution taking place. On the other hand, clearly distinct sister taxa such as humans and chimpanzees still share gene genealogy polymorphisms at some genes such as the HLA complex involved in immunological defence, and might therefore be classified as the same species under genealogical considerations.
C. Diagnostic species concept
The motivation for the diagnostic concept, usually called the "phylogenetic species concept" by its adherents, was again to incorporate phylogenetic Hennigian thinking into species-level taxonomy. There are many cases of hybridization between taxa on very different branches of species level phylogenies, which suggests that interbreeding and phylogeneticrealities conflict. Cracraft (1989) also noted that many bird taxa normally though of as subspecies were far more recognizable and stable nomenclaturally than the polytypic species to which they supposedly belonged (see also VIII D below). Cracraft therefore argued that the polytypic/interbreeding species concept should be rejected, and in its place we should use a diagnostic criterion in the form of at least one fixed difference at some inherited character. "A phylogenetic species is an irreducible (basal) cluster of organisms, diagnosably distinct from other such clusters, and within which there is a parental pattern of ancestry and descent" (Cracraft 1989). According to Cracraft, species defined in this way are the proper basal, real taxa suitable for phylogenetic analysis and evolutionary studies.
Of course, if diagnostic criteria are applied strictly, rather small groups of individuals, or even single specimens might be defined as separate species. Cracraft recognized this and argued that such diagnosable groups have no "parental pattern of ancestry and descent", i.e. they are not proper populations. However, this qualification appears similar to an interbreeding criterion of species, whereas the whole approach of using diagnostic characters was an attempt to get away from interbreeding.
Most evolutionary biologists balk at the idea of speciation being merely the acquisition of a new geographic diagnostic character, a DNA base pair or colour pattern change perhaps (Harrison 1998). "Speciation" is only a different, or special kind of evolution if the new "species" is a distinct population which can coexist locally with its sibling or parent population without losing its integrity.
Characters used to diagnose phylogenetic species may not be shared derived characters; they may be primitive (plesiomorphic) characters, or they may have evolved several times. Therefore, phylogenetic species need not be monophyletic, and could presumably be paraphyletic and perhaps polyphyletic. Cracraft appears confused on this matter: on the one hand he claims that phylogenetic species "will never be non-monophyletic, except through error" (Cracraft 1989: 35), but on the other recognizes that "their historical status may [sometimes] be unresolved because relative to their sister species they are primitive in all respects. Whether they ... [are] truly paraphyletic ... is probably unresolvable" (Cracraft 1989: 35). It seems odd to allow a "phylogenetic" species to be even paraphyletic (let alone polyphyletic) because paraphyly contravenes one of the basic tenets of phylogenetic systematics, and because one of the main justifications for a phylogenetic species concept is that species defined via other concepts might sometimes be paraphyletic: "The biological species concept cannot be applied to the Thomomys umbrinus complex unless one is willing to accept paraphyletic species, and do do so would be a de facto admission that biological species are not units of evolution" (Cracraft 1989: 46; see also Davis 1997: 374). The phylogeneticists' resolution of that problem, using diagnostic characters, leads to the same difficulty all over again! This rather glaring logical inconsistency considerably undercuts the argument for a diagnostic species concept.
In spite of these problems, Cracraft highlighted some genuine practical problems with the polytypic application of the interbreeding concept, and as a result this phylogenetic species concept has been influential. Ornithologists in particular have used diagnostic characters to reassign many taxa long thought of as subspecies to the level of full species. Recently many molecular systematists, including botanists (Davis 1997) have taken up Cracraft's suggestion and used diagnostic differences between populations, in some cases at single DNA base pairs, as evidence that two forms are separate species even if they intergrade freely at the boundaries of their distribution.
VII. Combined species concepts
A. Evolutionary and lineage concepts
Faced with the problem of studying the evolution of species through time, the palaeontologist G.G. Simpson (1951) proposed his "evolutionary species concept", in which a species is "a lineage (an ancestral-descendant sequence of populations) evolving separately from others and with its own unitary evolutionary role and tendencies". In other words, Simpson combined the idea that species were historical lineages with the concept of their evolutionary and ecological role. The key essence here appears to be "evolutionary independence". This concept appeals to phylogenetic systematists and palaeontologists alike, because of its historical dimension, and to neontologists because of its acknowledgement that biological mechanisms are what make the species real. De Queiroz (1998) is perhaps the most recent reviewer to propose that a single concept, which he calls "the general lineage concept", under which "species are segments of population-level lineages", underlies all other species concepts. According to De Queiroz, apparently competing species concepts merely emphasize different characters or criteria for species definition, but all acknowledge implicitly or explicitly that evolutionary separateness of lineage is the primary concept. This is a nice ideal, but evolutionary independence has little logical force in its application to actual forms that hybridize or undergo genetic exchange.
B. Cohesion concept of species
In similar vein, Templeton's (1998) "cohesion concept" fuses a number of competing ideas of species. Templeton argues that a combination of ecological and reproductive cohesion are important for maintaining a species' evolutionary unity and integrity, thereby incorporating components of the evolutionary, ecological, recognition, and interbreeding concepts. As well as applying to asexual taxa ("too little sex"), Templeton's idea also applied to species like oaks that undergo frequent hybridization and gene flow ("too much sex" for the interbreeding concepts). He further argues that separateness of genealogy is another important characteristic of species.
We are perhaps nearing the apogee of the species debate with these combined concepts. By incorporating evolutionary and phylogenetic origins together with every possible biological means by which species are currently maintained, these combined concepts "cover all the bases". One can acknowledge that species evolve and are maintained as cohesive wholes by all of these multifarious processes, yet at the same time argue that species can, and perhaps should be seen as separate from their histories of origin and from current reasons for their integrity. If groups with very different and conflicting biological and evolutionary characteristics are all considered species, there should exist a simpler criterion that unites them. It can also be argued that to conflate the origin and evolutionary role of a taxon with the definition of that taxon itself may lead to circularity, particularly in conservation or ecological studies, or when investigating speciation.
VIII. Dissent: maybe species are not real.
Throughout the history of the species debate, starting with Darwin, there have been some who argue that species are not individual "real" objects, but should instead be considered merely as man-made constructs merely useful in understanding of biodiversity and its evolution. These people are not necessarily nihilists, who deny that species exist: they simply argue that actual morphological and genetic gaps between populations would be more useful for delimiting species than inferred processes underlying evolution or maintenance of these gaps. By their refusal to unite these ideas under a single named concept, this biologist "silent majority" have rarely found a common voice.
A. Taxonomic practice
Taxonomists are on the front line of the species battle, because it is they who ultimately decide whether to lump or split taxa, and at what level to name them as species. If the objectivity and individuality of species as the primary taxon exists, taxonomists' activities have not been made any easier; and many taxonomists have simply ignored or denied belief in the evolutionary reality of species. In general it is probably true to say that at least 10% of taxonomic species are subject to revision because of these practical difficulties in delimitation.
For this reason, since the rise of the polytypic/interbreeding species concept, there has been little impact of the post-war species concepts on practising taxonomists, even while the debate raged around them. Procedure, at least in zoology, was more or less as follows: geographic variants which blended (or were thought to be able to blend) together at their boundaries were united within a single, polytypic species, unless morphological or genetic differences were so great that it seemed necessary to recognize two species. On the other hand, whenever two divergent forms differing at several unrelated traits, overlapped spatially, they were recognized as separate species even if a few intermediates suggested some hybridization or gene flow. Some taxonomists regarded subspecies as artificial taxa to be avoided, and may either have ignored geographic variation, or elevated subspecies of polytypic species to the rank of full species. But good taxonomic practice on species remained broadly similar across most branches of systematics, and involved careful analysis of multiple, chiefly morphological character sets tested in large samples of specimens collected from as many geographic regions as possible.
This view on species and subspecies has led in zoology to a steady reduction in the numbers of recognized species, as more and more dubiously separated taxa, previously ranked as species, became inserted as subspecies into larger and larger polytypic species. Recently, however, the diagnostic version of the phylogenetic species concept (VI C above) has been making strong inroads into zoological nomenclature, with the result is that counts of species on continents are again climbing as former subspecies are re-elevated to the species level, in spite of intergradation at their boundaries. However, the situation could get much worse; many Heliconius butterflies, for example, have over thirty geographic subspecies per species, all of which can be diagnosed easily. The numbers of bird and butterfly species could easily increase 2x - 10x in some groups if the diagnostic criterion were generally adopted. The one reality that is clear in species-level taxonomy is that the species is not real enough to remain at the same phylogenetic level while taxonomic fashions change. There is good evidence that actual species have been and still are purely man-made taxonomic units lacking in any objectively determinable underlying essence, even if such an essence exists.
B. Populations are evolutionary units, not species
Botanists deal with geographically variable organisms with low powers of dispersal, and have therefore never been very happy with the polytypic/interbreeding concept applied with such apparent success in zoology. Meanwhile, the strong surge in experimental population genetics and evolutionary studies that followed the books by Dobzhansky, Mayr and Stebbins has led to a greatly improved understanding of gene flow in natural populations. Gene flow in quite mobile animals, such as birds or butterflies, may not unite even adjacent local populations in a common gene pool. If local populations only rarely exchange genes, then gene flow across the range of a continental species is clearly insufficient to explain species integrity, because it would be outweighed easily by weak local patterns of adaptation or genetic drift (Endler 1977).
This increasing input of population biology into systematics and evolution led to the proposal by Ehrlich & Raven (1969), Levin (1979) and others that species are not real biological units at all; instead, local populations are the only real groupings united by gene flow within a common gene pool, and which adapt to local conditions, compete, and so on. Any homogeneity of ecological niche or genetics over the range of a species might be due either to simple evolutionary inertia or to similar stabilizing selection everywhere. To these authors species exist and are real in local communities; but it is fallacious to treat distant populations in the same way (see also VIII D below).
This viewpoint is generally understood and respected by population biologists, but curiously has not been incorporated explicitly into current thinking on species in systematics and evolution. Perhaps there is a sneaking suspicion that even very weak levels of gene flow may explain the species integrity over wide areas.
C. Phenetic species concept
In the 1960s and 1970s, a major systematics movement proposed numerical methods in taxonomy now usually referred to as phenetics. Pheneticists, as they were called, argued that taxonomy and systematics should be based on multivariate statistical analysis of characters rather than on underlying evolutionary or biological process information. If taxa were defined by non-evolutionary criteria, studies of evolution would be freed from the tautology of testing hypotheses about processes, when those same processes are used as assumptions in the definitions of taxa under study. Species, like other taxa, would be defined in numerical taxonomy on the basis of multivariate statistics, as clusters in phenotypic space (Sokal and Crovello 1970).
Phenetics is reviled by those who believe that classifications should be phylogenetic. However, the approach is closely similar to the intuitive methods adopted by most actual taxonomists, who use multiple morphological or genetic characteristics to sort individual specimens into discrete groups between which there are few intermediates (see V A above). Some large areas of practical taxonomy are based purely on this phenetic approach. Bacterial systematists, for instance, use multiple biochemical tests to assign microbes to species taxa. The usefulness of this taxonomic method is attested by its success in hospitals for predicting pathogeneticity and antibiotic sensitivity.
Phenetic classifications based on morphology introduce the danger that, if convergent characters are used as data, one may group unrelated forms into paraphyletic or even polyphyletic taxa. In addition, single gene polymorphisms and sexual dimorphism can affect multiple morphological characters. This could lead to recognition of multiple species within polymorphic populations. Sibling species, on the other hand, could be lumped into the same species using a phenetic approach, unless a set of highly diagnostic characters could be found.
D. Genotypic cluster criterion
For morphological or genetic gaps to exist between species, gene flow between species (if any) must be balanced by an opposing force of disruptive selection. In my own work, I had studied hybrid zones between geographic forms of butterflies, and I attempted to show that a practical statistical definition of species could be constructed using morphological and genetic gaps alone, rather than using phylogetic or evolutionary processes that caused the gaps to exist.
However, to define species by means of the gaps between them requires consideration of the nature of the gaps to avoid falling into the trap of defining polymorphic forms as separate species, or of lumping sibling species. Rather than merely using external morphology, in difficult cases I proposed that we could consider the genetics as well. DNA has a digital, rather than analogue code, so there is a genetic gap between virtually any pair of individuals. Clearly, then, we cannot use just any discreteness at the genetic level to define species. Separate sexes and polymorphic female forms of mimetic Papilio butterflies also have gaps between them in exactly this way. A genetic element, which may be a single base pair, an allele at a gene, the entire mitochondrial genome, a chromosomal rearrangement, or perhaps a sex chromosome, may determine the genetic or morphological differences between such polymorphic forms.
To be considered part of a single local population, and therefore part of the same local species, we expect that polymorphic genetic elements like mimicry genes and sex chromosomes will be mixed in with polymorphisms at genetic elements found on other chromosomes or extrachromosomal DNA. Each individual may be a distinct multilocus genotype, but we recognize groupings of these genotypes because a polymorphism at one genetic element is more or less independent of polymorphisms at another. Conversely, if alleles at one locus are strongly associated with alleles at other, unlinked elements, we have evidence for two separate populations or species.
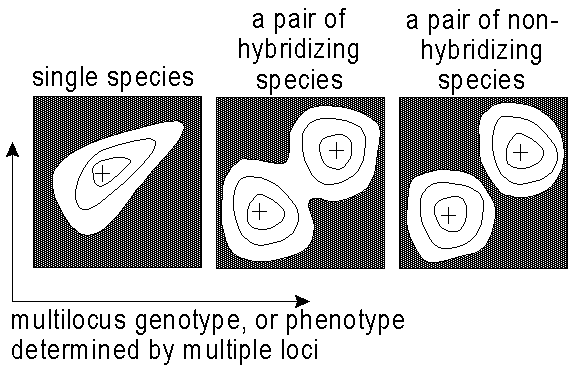
I therefore proposed a "genotypic cluster criterion" of species. Species are recognized by morphological and genetic gaps between populations in a local area rather than by means of the phylogeny, cohesion, or reproductive isolation that are responsible for these gaps (Mallet 1995). In a local area, a single species (the null hypothesis) is recognized if there is but a single cluster in the frequency distribution of multilocus phenotypes and genotypes. Separate species are recognized if there are several clusters separated by multilocus phenotypic or genotypic gaps. These gaps may be entirely vacant, or they may contain low frequencies of intermediate genotypes, or hybrids (Fig. 1). The definition is useful because one avoids tautological thinking: hypotheses about speciation or phylogeny become independent of assumptions about the nature of reproductive isolation or phylogeny underlying the taxa studied.
Fig. 1. Genotypic cluster criterion for species. A sample of individuals is made at a single place and time. Numbers of individuals are represented by the contours in multidimensional genotypic space. Peaks in the abundance are represented by +. Two species are detected if there are two peaks in these distributions (right, bimodal distribution), otherwise the null hypothesis of a single species is not rejected (left, unimodal distribution). Note: the axes represent multidimensional morphological/genotypic space, not geographic space.
Genotypic clusters are neither profound nor original; I trace the
idea to Darwin (1859, see IIIA), although it undoubtedly goes further back
since its use does not require acceptance of evolution. Many similar proposals
have been made (Simpson 1937, Hutchinson 1968, Sokal & Crovello 1970,
Avise & Ball 1990, Cohan 1994, Smith 1994); in fact the approach is
similar to the practice used on morphology by most taxonomists (see VIIIA),
the phenetic concept (see VIIIC) and indeed by those who attempt a practical
application of the biological species concept (see below). This widespread
use of direct genotypic or morphological data, as opposed to reproductive
or phylogenetic inferences made from such data, has apparently lacked support
because its practitioners were loath to label it as a "concept". I intended
merely to justify the Darwinian and practical taxonomists' species definition
statistically and in terms of genetics, rather than to enforce the use
of genotypes instead of morphology to define species. Most genotypic cluster
species can be recognized morphologically; for example, minor pattern elements
in Papilio can be used to unite the various polymorphic forms; however,
with abundant molecular marker data we could easily use the criterion to
sort actual specimens.
There is also nothing wrong with concluding after seeing a male butterfly mating with an unlike female that they belong to the same species, but, because hybridization does occur occasionally between forms normally thought of as different species, one is not so much using the mating behaviour itself to define species as inferring that the mating behaviour is a common event. We believe that, if we were to analyse their genomes, the two hybridizing forms would have similar genetic characteristics apart from those determining sexual dimorphism, i.e. they would belong to the same genotypic cluster. Instead of reproductive compatibility being the primary criterion of species, we can turn the argument on its head: we infer from limited data on reproductive compatibility that a single genotypic cluster may result.
Asexual forms, unclassifiable under the interbreeding concept, and arbitrarily definable at any level under concepts depending on phylogeny, can be clustered and classified as a genotypic clusters in exactly the same way as sexual species. The precise taxonomic level of species clustering is potentially somewhat arbitrary, as in the phylogenetic concepts, but at least the method acknowledges this arbitrariness rather than pretending that some higher evolutionary principle is being used. However, many asexual forms such as bdelloid rotifers have easily distinguishable species taxa (Hutchinson 1968). In bacteria, competition is thought to structure promiscuous, but largely asexual populations into recognizable genetic clusters (Cohan 1994, see also V A above).
Critics have argued that the genotypic cluster criterion in sexual species is nothing other than a gene flow concept of species under a different guise. This is true for one specialized interpretation of "gene flow". If we define gene flow as the successful or effective as opposed to actual input of genes, then it is easy to see a "gene flow criterion" becomes similar to the "genotypic cluster" criterion: to find whether a hybridization or gene flow event is successful, we must either follow every gene through all possible descendants for all time, or we may examine the genotypic state of a population and determine if genes from one form are mixed randomly with genes from another form. It seems clear that looking for random association of genes within genotypes in the genotypic cluster approach will be methodologically the same as genotypic state analysis to determine whether a population is interbreeding, but the latter requires additional assumptions. The genotypic cluster criterion in sexual species could be looked upon simply as a practical application of the biological species concept. However, I distinguish the genotypic cluster criterion from the interbreeding concept, if only because its name emphasizes that the definition is character-based, rather than actually based on interbreeding, and is thus applicable to asexuals as well as sexual species.
If a single geographic race, which previously intergraded at all its boundaries with other geographic races, were to split into two forms that coexist as separate genotypic clusters, we could have a situation that the original polytypic species became paraphyletic. The new species has been derived from only one of the component subspecies. Thus paraphyly of species must be recognized as a possibility under this definition, as with both interbreeding and diagnostic concepts.
E. The unreality of species in space and time
Geographic races often form clusters differing at multiple loci from other races in the same species. The interbreeding concept or genotypic cluster criterion can be used to justify a classical polytypic species if the various geographic races are separated by zones which contain abundant intermediates (hybrids). We sample multilocus genotypes or phenotypes in local areas of overlap and determine whether a single peak (one species, i.e. abundant hybrids), or two peaks (two different species, i.e. rare hybrids) are evident in the local genotypic distribution (Fig. 1). Hybridization may occur, but if it is rare so that character and genotypic distributions remain distinctly bimodal in zones of overlap, we usually classify them as separate species, even under the interbreeding concept.
Although this spatial extension of the local species is practical to apply to any pair of forms in contact, it is unlikely to lead to general agreement. The problem is that hybrid zones can be very narrow and may separate forms that are highly distinct at multiple characters or loci, in spite of complete unimodal blending in local areas of overlap. Even adherents of the interbreeding concept are reluctant to lump such geographic forms within the same species. Examples include North American swallowtail butterflies (Papilio glaucus/P. canadensis, see Hagen et al. 1991) and European toads (Bombina bombina/B. variegata, see Szymura 1993).
An even worse problem is found in "ring species", which form a continuous band of intergrading subspecies, but whose terminal taxa may be incompatible, and overlap without intergrading. A commonly cited example are the herring gulls and lesser black-backed gulls (Larus argentatus complex, Mayr 1970). Similarly, while most hybrid zones between European Bombina are unimodal, the same pair of taxa may have bimodal genotypic distributions in other zones of overlap (Szymura 1993). Thus, geographic forms may be apparently conspecific in some areas, but overlap as separate species in other areas. Finally, if distinct populations are geographically isolated and there is no area of overlap, one cannot disprove the null hypothesis of "same species" under interbreeding or genotypic cluster criteria, but biologists are reluctant to unite such populations if they are very divergent. Laboratory hybridization could be tried, but many overlapping species are known to hybridize freely in captivity, while remaining separate in nature. There are good examples in the great apes, for instance the bonobo (Pan paniscus) versus the chimpanzee (P. troglodytes), and the gorillas (Uchida 1996), but similar decisions must be made in almost any animal and plant group.
The problem for spatial extensions of species criteria is in fact due to the evolutionary divergence of spatially separated lineages: time and space are correlated. Paleontologists face a similar temporal problem when classifying fossils in different strata. Evolutionary rates may vary, but all lineages must ultimately be continuous, so there is no very logical place to put a species boundary in time any more than there is in space. Paleontologists typically use operational species on the basis of morphological gaps between taxa from the same and different time periods (Simpson 1937, Smith 1994).
These difficulties show that there is no easy way to tell whether related geographic or temporal forms belong to the same or different species. Species gaps can be verified only locally and at a single time. One is forced to admit that Darwin's insight is correct: any local reality or integrity of species is greatly reduced over large geographic ranges and time periods (see also Mayr 1970, and IX D).
IX. The importance of species concepts for biodiversity and conservation
A. Traditional: species as real entities
Different species concepts seek to define species in mutually incompatible ways. Thus, a monophyletic species concept seems not very useful to evolutionary biologists because of difficulties with multiple gene genealogies and paraphyletic remnants. In contrast, the interbreeding concept and other combined concepts incorporating biological processes of species maintenance suffer in the eyes of phylogenetic systematists because they lack phylogenetic coherence and produce paraphyletic taxa, or worse. If we were to allow the basal unit of our taxonomy to incorporate paraphyly, it would be harder to justify a strict adherence to monophyly at other taxonomic levels. It is beyond the scope of this article to resolve these difficult issues, but these conceptal conflicts fuel the continuation of the debate, and also highlight the fact that if species are indeed real, objective biological units, their unifying reality has been extremely difficult to verify.
Many ecological and biodiversity studies of actual organisms ignore these difficulties, and assume that species are objectively real basal units. Thus in ecology, we have theories of global species diversity. In conservation, we have the "Endangered Species Act" in the USA, which prescribes the conservation of threatened taxa we call species. Populations not viewed as species, particularly putative hybrid taxa (like the red wolf, Canis rufus, of the SE USA), are seen as less valuable, even if rare. How do we recognize that a taxon is hybridized? Obviously, to be a hybrid, it must be a mere intergrade between two, real, objectively identifiable entities. The Endangered Species Act views species as important real conservation units and hybrids as unimportant. It did this because it incorporated the species concept in vogue at the time of its enactment, i.e. the biological species concept, in which hybridization is seen as a "breakdown in isolating mechanisms" (Mayr 1970).
B. Alternatives: genetic differences more valuable than species status
If this Act were to be rewritten today, what would it say? There is undoubtedly a greater realization today that other levels in the taxonomic hierarchy are important elements of biodiversity. The diagnostic concept of species, while claiming to support the basal, objective nature of species, can at least have the beneficial effect of allowing its basal unit of biodiversity to be recognized at a lower level, in this case as subspecies within polytypic species. Some molecular geneticists have advocated conservation of "evolutionary significant units", "management units", or "stocks" (a fisheries term) defined on criteria of continuous genetic differentiation at molecular markers (Moritz 1994) as being more important than worrying about the species level at all. The true reality of spatiotemporally extended species eludes us, if it exists. If this is so then it seems best to adopt some other measure of conservation value that relies purely on the degree of genetic differentiation, for instance at molecular genetic markers.
C. Species differences as ecologically important markers.
However, there are many who oppose this view. Species within a local area such as a nature reserve are, for the most part, easily and objectively identifiable using morphology, behaviour, genetics, or phylogeny. A pair of similar species must usually be ecologically different to coexist. To remain distinct, sexual species will need some prezygotic isolation, so their mating behaviour must also be different. Thus, counting species in a local area makes ecological sense, and conserving species diversity in a local area would conserve actual ecological and behavioural diversity. Behavioural and morphological differences that cause species-ness are evolutionarily more valuable than potentially neutral genetic differences at molecular markers.
D. Biodiversity in space and time.
As we have seen, this local view breaks down when we try to apply the term "species" over large areas or geological time scales. In some cases, there is excellent homogoneity over large areas; for example the painted lady butterfly (Vanessa cardui) and the barn owl (Tyto alba) have a virtually worldwide distribution and look nearly identical everywhere. Other species are not so homogeneous: the familiar mallard group of ducks (Anas platyrhynchos) has become highly differentiated into some 18 or so forms in far-flung outposts of the world, but exactly how many are good species, and how many are races, or indeed, how many races there are in total, is a matter of taste. Current authorities recognize about 10 species, but there might easily be five or 15 in alternative treatments. One of the forms, the Mexican duck A. platyrhynchos diazi is threatened with hybridization by its mallard relative Anas platyrhynchos platyrhynchos which has been expanding from the north, and the American black duck (Anas rubripes) also hybridizes with the mallard, but appears to resist hybridization somewhat better than the Mexican form - hence its species status.
Faced with these difficulties, should we worry about the species level when conserving endangered taxa over large areas? Whatever the answer to this question, it does not seem very sensible to rely too much on the spatiotemporal reality of species for an answer. We might rename the Mexican Duck as a separate species instead of a subspecies, but ideally this should not affect its conservation value since there has been no actual change in the knowledge of biological characteristics that affect conservation value. Most conservationists now agree that the former fetish for species-level legislation was a mistake: conservation and legislation should now recognize that living, evolving populations form fractal continua over time and space, rather than attempting a division into spurious "fundamental" units.
Species are certainly fundamental units of local biodiversity, but they have this clarity only in a small zone of time and space, and so species counts become less and less meaningful as larger and larger areas are covered. Taxonomists might come to nominalistic agreements on a case-by-case basis, but even this shows little sign of happening yet. Ecological theory, as well as conservation and biodiversity studies must recognize that species counts over large expanses of space and time represent only a sketchy measure of biodiversity, a measure which owes more to taxonomic and metaphysical fashion than to science.
GLOSSARY
Cladistic a classification based entirely on monophyletic taxonomic
groupings within a phylogeny; taxonomic units that are paraphyletic or
polyphyletic are rejected. A cladist is one who practices cladistics, usually
in the sense of using parsimony to adjudicate between data from multiple
characters in the construction of a cladogram, which is an estimate of
the true phylogeny.
Cohesion the sum total of forces or systems that hold a species together.
The term is used especially in the interbreeding and cohesion species concepts.
Cohesion mechanisms include isolating mechanisms in sexual species as well
as stabilizing ecological selection, which may cause cohesion even within
asexual lineages.
Disruptive selection selection acting to preserve extreme phenotypes
in a population. Speciation usually involves disruptive selection, because
intermediates (hybrids between incipient species) are disfavoured (see
also stabilizing selection).
Gene flow movement of genes between populations, usually via immigration
and mating of whole genotypes, but sometimes single genes may undergo horizontal
gene transfer via transfection by microorganisms.
Gene pool the sum total of the genetic variation within a reproductively
isolated species population; this term is mostly used by supporters of
the interbreeding species concept.
Isolating mechanisms the sum total of all types of factors that prevent
gene flow between species, including pre-mating mechanisms (mate choice),
and post-mating mechanisms (hybrid sterility and inviability). Modern authors
deny that these "mechanisms" have necessarily evolved to preserve the species'
integrity as originally assumed, though this may sometimes be the case
in reinforcement of pre-mating isolation. Isolating mechanisms are a subset
of the factors that cause cohesion of species under the interbreeding and
cohesion species concepts.
Monophyletic a grouping that contains all of the descendants of a particular
node in a phylogeny. Monophyly is the state of such groupings. Compare
paraphyletic, polyphyletic. Butterflies (Rhopalocera) and birds (Aves)
are examples of two groups thought to be monophyletic.
Paraphyletic a grouping that contains some, but not all of the descendants
of a particular node in a phylogeny. Paraphyly is the state of such groupings.
Compare monophyletic, polyphyletic. Moths (Lepidoptera, excluding butterflies)
and reptiles (amniotes, excluding birds and mammals) are examples of two
groups thought to be paraphyletic.
Phenetic, a classification or grouping based purely on overall similarity.
Pheneticists use matrices of overall similarity rather than parsimony to
construct a phenogram as an estimate of the phylogeny. Examples of phenetic
methods of estimation include unweighted pair group analysis (UPGMA) and
neighbor joining. Cladists reject phenetic classifications on the grounds
that they may result in paraphyletic or polyphyletic groupings.
Phylogenetic pertaining to the true (i.e. evolutionary) pattern of
relationship, usually expressed in the form of a binary branching tree,
or phylogeny. If hybridization produces new lineages, as is common in many
plants and some animals, the phylogeny is said to be "reticulate". Phylogenies
may be estimated using phenetics, parsimony (cladistics) or methods based
on statistical likelihood.
Polyphyletic groupings contain taxa with more than one ancestor. Polyphyly
is the state of such groupings. Compare paraphyletic and monophyletic.
"Winged vertebrates" (including birds and bats) give an example of a polyphyletic
group.
Real, reality two tricky words found frequently in the species concept
debate. "Reality" is typically used to support one's own species concept,
as in: "The conclusions set forth above ... lead to a belief in the reality
of species" (Poulton 1904); similar examples can by found in the writings
of Dobzhansky, Mayr, and especially phylogeneticists. The term reality
in this sense is similar to an Aristotelian "essence", a hypothetical pure,
albeit obscure truth that underlies the messy actuality; unfortunately,
in everyday language "real" also means "actual" (curiously, a "reality"
in the first sense may be "unreal" under the second!). By rejecting the
"reality" of species, one can therefore send very mixed messages: some
readers will understand the author to be a nominalist who merely believes
useful terms require little theoretical underpinning; others assume the
author is nonsensically using some definition that does not apply to actual
organisms. Here, when I discuss the reality underlying a species concept,
I mean it in the first sense, a hypothesis. Many authors of species concepts
and some philosophers of science argue that definitions must be underpinned
by a theoretical justification or reality. Other philosophers such as Wittgenstein
and Popper agree that terms need no such definition to be useful.
Speciation the evolutionary process of the origin of a new species.
Specific mate recognition systems (SMRS) fertilization and mate recognition
systems in the recognition concept of species, the factors leading to pre-mating
compatibility within a species. See also cohesion which is similar to SMRS,
but includes post-mating compatibility as well.
Sibling species a pair of closely related, morphologically similar
species (usually sister species).
Stabilizing selection selection which favours intermediate phenotypes.
BIBLIOGRAPHY
Avise, J. C., and R. M. Ball. 1990. Principles of genealogical concordance in species concepts and biological taxonomy. Pp. 45-67 in D. J. Futuyma and J. Antonovics, eds. Oxford Surveys in Evolutionary Biology, vol. 7. Oxford Univ. Press, Oxford.
Baum, D. A., and K. L. Shaw. 1995. Genealogical perspectives on the species problem. Pp. 289-303 in P. C. Hoch and A. G. Stephenson, eds. Experimental and Molecular Approaches to Plant Biosystematics. Missouri Botanical Garden, St. Louis, Missouri. (Monographs in Systematic Botany from the Missouri Botanical Garden; 53).
Claridge, M. F., H. A. Dawah, and M. R. Wilson, eds. 1997. Species: The Units of Biodiversity. Chapman & Hall, London.
Cohan, F. M. 1994. The effects of rare but promiscuous genetic exchange on evolutionary divergence in prokaryotes. Amer. Nat. 143:965-986.
Cracraft, J. 1989. Speciation and its ontology: the empirical consequences of alternative species concepts for understanding patterns and processes of differentiation. Pp. 28-59 in D. Otte and J. A. Endler, eds. Speciation and its Consequences. Sinauer Associates, Sunderland, Mass.
Darwin, C. 1859. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, 1st ed. John Murray, London.
Davis, J. I. 1997. Evolution, evidence, and the role of species concepts in systematics. Syst. Bot. 22:373-403.
de Queiroz, K. 1998. The general lineage concept of species, species criteria, and the process of speciation. A conceptual unification and terminological recommendations. Pp. 57-75 in D. J. Howard and S. H. Berlocher, eds. Endless Forms. Species and Speciation. Oxford University Press, New York.
Ehrlich, P. R., and P. H. Raven. 1969. Differentiation of populations. Science 165:1228-1232.
Endler, J. A. 1977. Geographic Variation, Speciation, and Clines. Princeton University Press, Princeton, N.J.
Hagen, R. H., R. C. Lederhouse, J. L. Bossart, and J. M. Scriber. 1991. Papilio canadensis and P. glaucus (Papilionidae) are distinct species. J. Lepid. Soc. 45:245-258.
Harrison, R. G. 1998. Linking evolutionary pattern and process. The relevance of species concepts for the study of speciation. Pp. 19-31 in D. J. Howard and S. H. Berlocher, eds. Endless Forms. Species and Speciation. Oxford Univ. Press, New York.
Hennig, W. 1966. Phylogenetic Systematics. University of Illinois Pres, Urbana.
Howard, D. J., and S. H. Berlocher, eds. 1998. Endless Forms. Species and Speciation. Oxford Univ. Press, New York.
Hutchinson, G. E. 1968. When are species necessary? Pp. 177-186 in R. C. Lewontin, ed. Population Biology and Evolution. Syracuse University Press, Syracuse, NY. Proceedings of the International Symposium Sponsored by Syracuse University and the New York State Science and Technology Foundation. June 7-9, 1967, Syracuse, New York.
Levin, D. A. 1979. The nature of plant species. Science 204:381-384.
Maddison, W. P. 1997. Gene trees in species trees. Syst. Biol. 46:523-536.
Mallet, J. 1995. A species definition for the Modern Synthesis. Trends Ecol. Evol. 10:294-299.
Mayr, E. 1970. Populations, Species, and Evolution. Harvard University Press, Cambridge, Mass.
---. 1982. The Growth of Biological Thought. Diversity, Evolution, and Inheritance. Belknap, Cambridge, Mass.
Moritz, C. 1994. Defining 'Evolutionarily Significant Units' for conservation. Trends Ecol. Evol. 9:373-375.
Paterson, H. E. H. 1985. The recognition concept of species. Pp. 21-29 in E. S. Vrba, ed. Species and Speciation. Transvaal Museum, Pretoria. (Transvaal Museum Monograph; 4).
Poulton, E. B. 1904. What is a species? Proc. Entomol. Soc. Lond. 1903:lxxvii-cxvi.
Ridley, M. 1996. Evolution. Blackwell Science, Oxford.
Robson, G. C. 1928. The Species Problem. An Introduction to the Study of Evolutionary Divergence in Natural Populations. Oliver and Boyd, Edinburgh.
Simpson, G. G. 1937. Patterns of phyletic evolution. Bull. Geol. Soc. Amer. 48:303-314.
---. 1951. The species concept. Evolution 5:285-298.
Smith, A. B. 1994. Systematics and the Fossil Record. Documenting Evolutionary Patterns. Blackwell Scientific, Oxford.
Sokal, R. R., and T. J. Crovello. 1970. The biological species concept: a critical evaluation. Amer. Nat. 104:107-123.
Szymura, J. M. 1993. Analysis of hybrid zones with Bombina. Pp. 261-289 in R. G. Harrison, ed. Hybrid Zones and the Evolutionary Process. Oxford University Press, New York.
Templeton, A. R. 1998. Species and speciation. Geography, population structure, ecology, and gene trees. Pp. 32-43 in D. J. Howard and S. H. Berlocher, eds. Endless Forms. Species and Speciation. Oxford Univ. Press, New York.
Uchida, A. 1996. What we don't know about great ape variation. Trends Ecol. Evol. 11:163-167.
Wallace, A. R. 1865. On the phenomena of variation and geographical distribution as illustrated by the Papilionidae of the Malayan region. Trans. Linn. Soc. Lond. 25:1-71.